


Cart (0 Items)
Your cart is currently empty.
View Products Blog
Blog
 Claire Perrot
Claire Perrot
This article breaks down the key difference between humanized and fully human antibodies: “mostly human” vs. “entirely human.” It compares leading discovery methods like transgenic mice, single B cell tech, phage display, and CDR grafting, showing when each makes the most sense. The goal: help researchers choose the smartest path for their therapeutic antibody projects.
Antibodies are proteins designed to recognize and neutralize specific targets (antigens). But when antibodies are derived from non-human species, such as mice, they can trigger unwanted immune responses in patients.

Humanized antibodies:
Fully human antibodies:
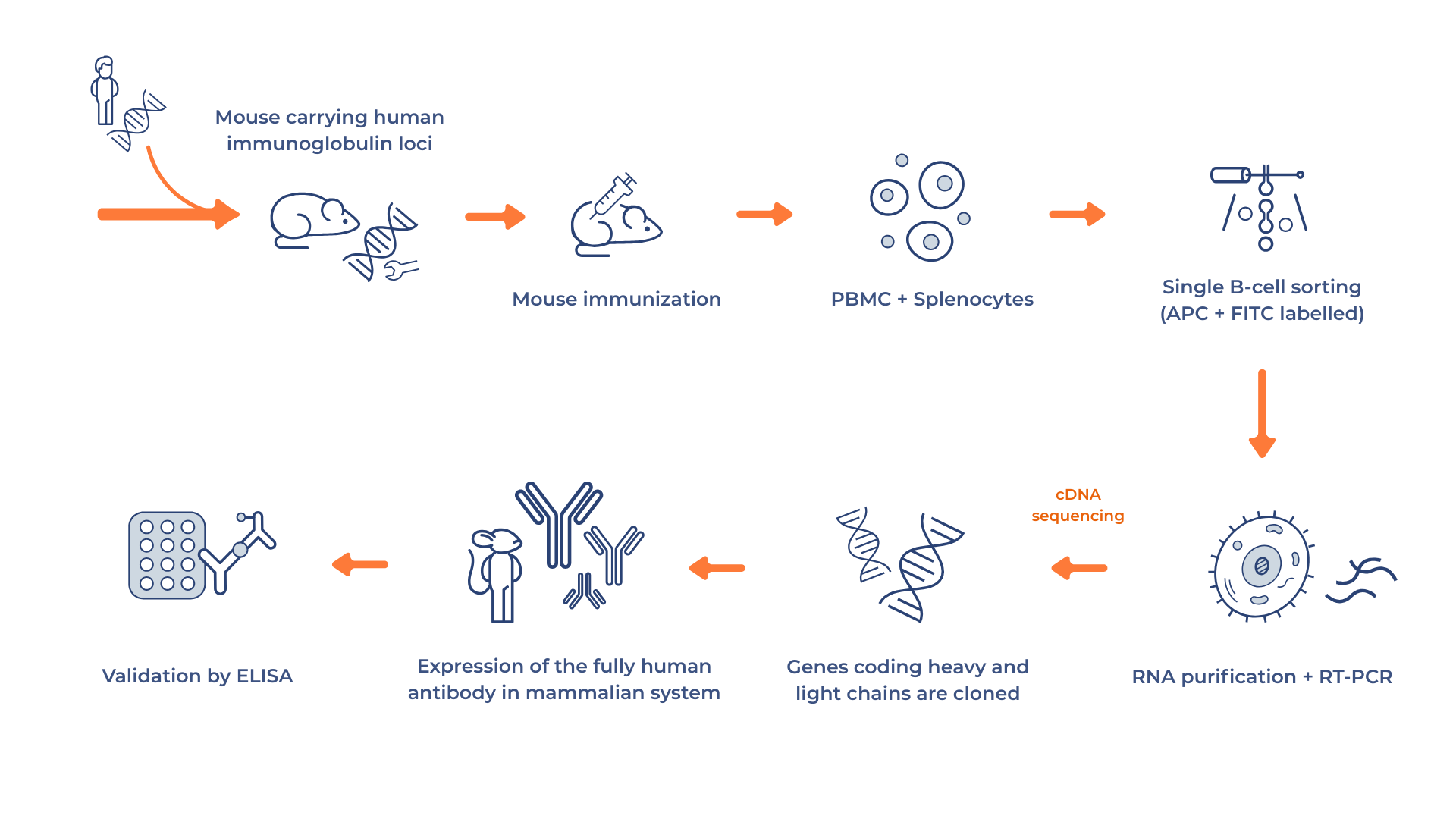
Humanized mice are genetically engineered to carry human immunoglobulin gene loci. After being immunized with a target antigen, they produce fully human antibodies that are harvested. Transgenic mice are genetically engineered to carry human immunoglobulin gene loci. After being immunized with a target antigen, they generate human B cells that can be harvested for antibody discovery. These B cells, can then be screened at the single-cell level to recover antibodies targeting clinically relevant epitopes with precision.
Advantages:
The key advantage of this approach is that the antibodies benefit from natural in vivo affinity maturation, which results in molecules with high specificity and favorable biophysical properties, closely resembling those of approved drugs. Because the antibodies are entirely human in sequence, the risk of immunogenicity is very low, making them well suited for therapeutic use. Another strong benefit is that the process does not require a separate humanization step, saving valuable time and effort in development. It is no coincidence that many blockbuster therapeutics, including Humira®, Dupixent®, Repatha®, and Praluent®, were generated using this method.
Limitations:
However, the limitations should not be overlooked. The reliance on animals introduces ethical and regulatory considerations that require specialized infrastructure, time, and approvals, often making the process slower and more expensive compared to in vitro alternatives. Single B cell workflows also require advanced instrumentation, bioinformatics, and highly precise handling. Additionally, this approach is less effective for developing antibodies against self-antigens or highly conserved targets, where the immune response of the mouse may be limited.
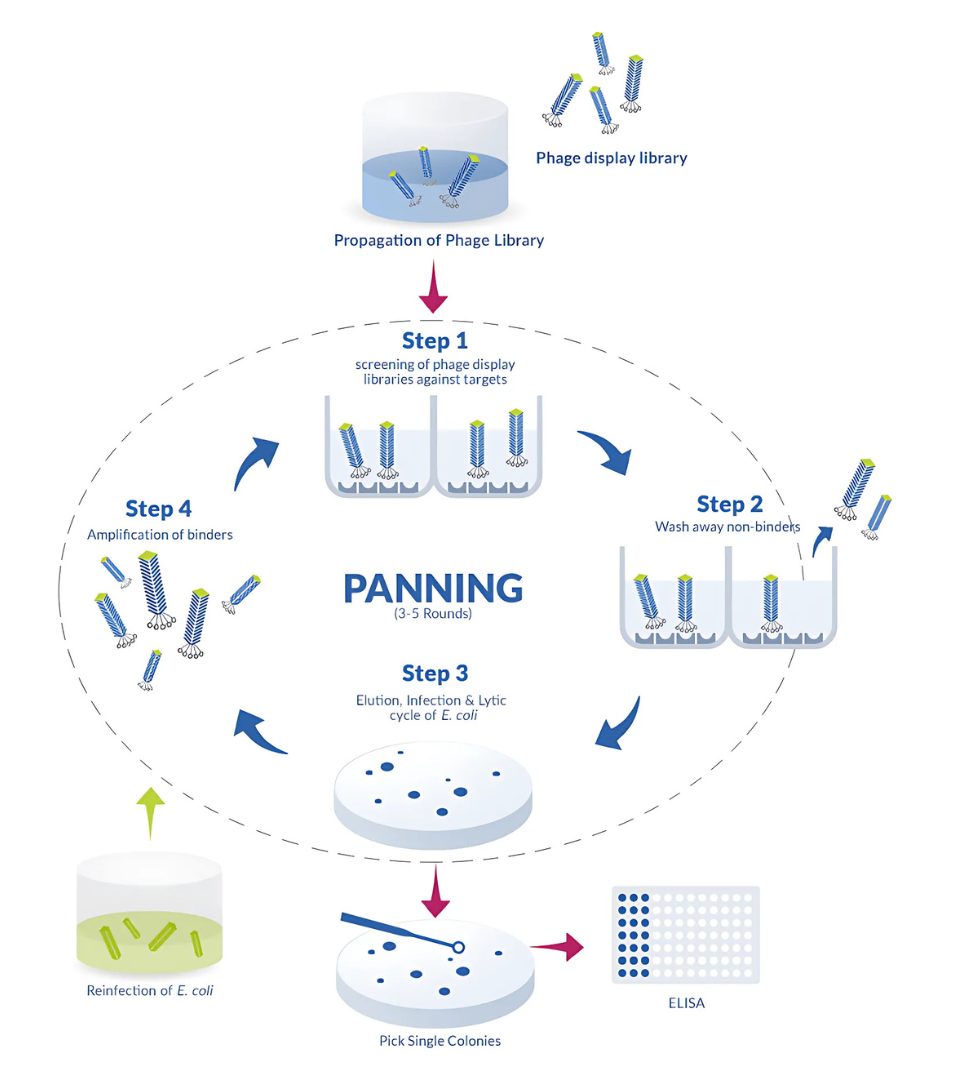
Phage display is an entirely in vitro method that relies on expressing human antibody gene fragments on the surface of bacteriophages. High-affinity binders are then isolated through a process known as biopanning, which enables screening from very large and diverse libraries that can reach up to 10¹¹ variants.
Advantages:
This technique offers significant advantages, particularly in terms of speed and scalability. Because it bypasses the need for immunization or animal models, researchers can rapidly identify binders against virtually any target, including toxic molecules or antigens that fail to trigger a strong immune response in vivo. This makes phage display an especially valuable tool for urgent applications, such as rapid lead generation during outbreak responses.
Limitations:
Yet, the method also comes with limitations. Since it lacks the natural affinity maturation provided by an immune system, additional engineering steps are often required to optimize the binders. The antibodies recovered are frequently in fragment formats such as scFvs, which must be converted into full-length molecules before therapeutic use. Although in vitro maturation strategies exist, they add complexity and time to the process.
ProteoGenix goes one step further by offering the world’s first cancer-focused human antibody library, specifically designed to accelerate the discovery of next-generation therapeutics.
cancer antibody discovery service
Beyond human repertoires, phage display can also be performed with non-human libraries, including murine, camelid, and synthetic antibody repertoires. These systems broaden the structural diversity of the antibodies and can provide unique properties that are difficult to obtain with human sequences alone.
Advantages:
The main advantage of non-human libraries is their ability to expand the discovery space and generate antibodies with novel frameworks. Camelid-derived single-domain antibodies, also known as NANOBODY® proteins*, are exceptionally small and stable, giving them access to cryptic epitopes that conventional antibodies cannot reach. Synthetic libraries further enhance this potential by introducing targeted diversity into specific regions of the antibody, allowing for tailored solutions against particularly challenging antigens. This makes non-human libraries a powerful tool when innovative formats or unusual binding properties are required.
Limitations:
Despite their strengths, these approaches still carry limitations. Because the sequences are non-human in origin, antibodies derived from these libraries usually require a subsequent humanization step to reduce immunogenicity, adding time and complexity to the workflow. In addition, while synthetic libraries offer extensive diversity, they cannot fully replicate the natural pairing and maturation processes of an immune system, which sometimes results in binders that need further optimization to improve affinity, stability, or overall developability.
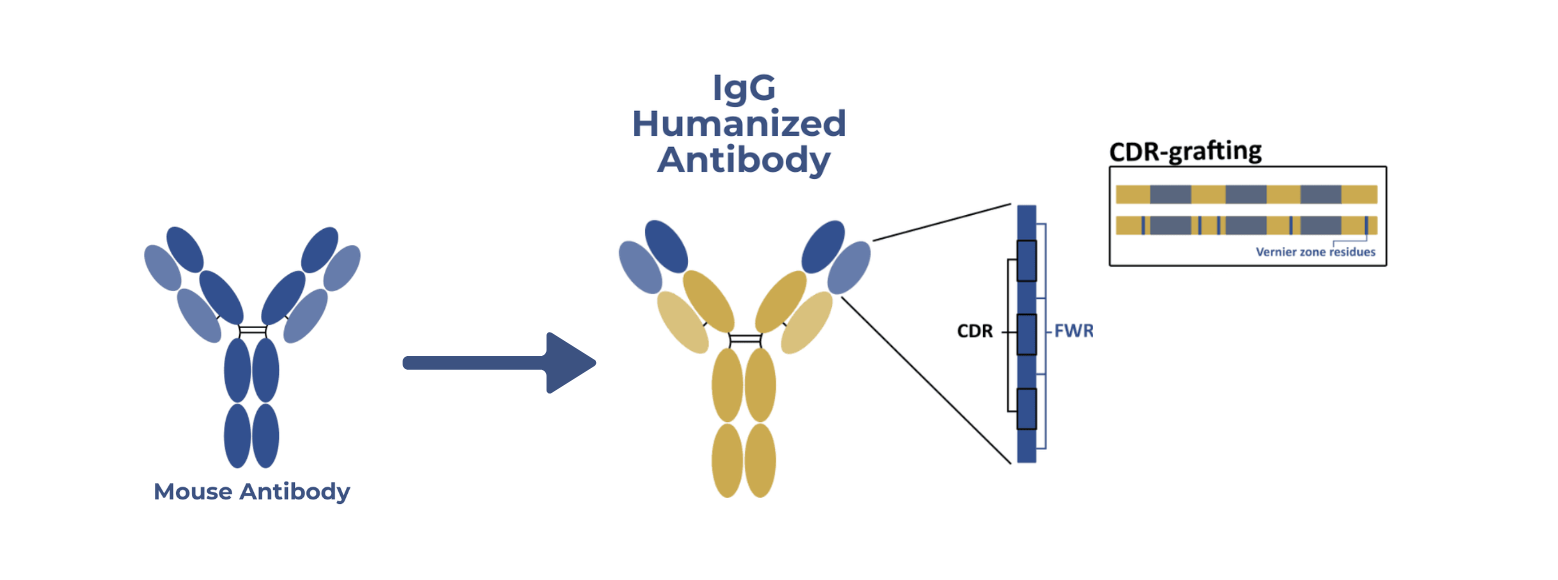
When a murine antibody already exists, humanization can be performed by grafting the antigen-binding complementarity-determining regions (CDRs) onto a human antibody framework.
Advantages:
The benefit of this strategy lies in its ability to preserve antigen specificity while simultaneously lowering the risk of immunogenicity, since the resulting molecule can often achieve more than 90% human sequence identity. It is particularly valuable when researchers are working with legacy murine antibodies or validated preclinical leads that have already demonstrated promising results.
Limitations:
The approach, however, is not without drawbacks. The grafting process can sometimes lead to a loss of affinity or functional activity, requiring extensive back-mutations or structural optimization to restore performance. Furthermore, because the CDRs remain murine in origin, there is still a residual risk that patients may develop anti-drug antibodies, which can impact safety or efficacy during clinical use.
Your choice depends on several factors:
| Method | Human Content | Speed | Cost | Best Use Case | Advantages | Limitations |
|---|---|---|---|---|---|---|
| Transgenic Mice | 100% | Moderate | High | Clinically validated therapeutics | Natural affinity maturation, drug-like props | Animal use, higher cost, slower timelines |
| Single B Cell | 100% | Moderate-Fast | High | Infectious disease, personalization | Truly human, natural pairing, relevant epitopes | Technically complex, lower diversity |
| Phage Display (Human) | 100% | Fast | Moderate | Rapid lead generation | Scalable, no animals, diverse repertoires | No natural maturation, fragment formats |
| Phage Display (Non-Human) coupled with humanization |
~90–95% | Fast | Moderate | Novel formats, hard targets | Unique scaffolds, NANOBODY® protein*, synthetic design | Needs humanization, less natural pairing |
| CDR Grafting (Humanization) | ~90–95% | Moderate | Moderate | Legacy murine leads | Preserves specificity, lowers immunogenicity | Risk of affinity loss, residual murine CDRs |
At ProteoGenix, we support every stage of antibody generation:
Our expertise allows us to tailor the best strategy to your antigen, therapeutic goals, and timeline. Importantly, we take into account the inherent limitations of each method, whether that means optimizing affinity after CDR grafting, complementing phage display with in vitro maturation, or implementing streamlined processes to reduce the timelines and costs of animal-based approaches. This way, we ensure your project benefits from the strengths of each technology while minimizing potential drawbacks.
Contact our team to discuss your antibody project
*NANOBODY® compound is a registered trademark of Ablynx N.V.
Almagro, J. C., Pedraza-Escalona, M., Arrieta, H. I., & Pérez-Tapia, S. M. (2019). Phage display libraries for antibody therapeutic discovery and development. Journal of Biomedical Science, 26, Article 31. https://doi.org/10.1186/s12929-019-0592-z
Carter, P. J., & Lazar, G. A. (2021). Next generation antibody drugs: Pursuit of the “high-hanging fruit.” Nature Reviews Drug Discovery, 20(5), 354–355. https://doi.org/10.1038/s41573-021-00175-8
Chen, H., et al. (2022). AI in antibody discovery. Frontiers in Bioinformatics, 2, Article 1044975. https://doi.org/10.3389/fbinf.2022.1044975
Ecker, D. M., Jones, S. D., & Levine, H. L. (2015). The therapeutic monoclonal antibody market. mAbs, 7(1), 9–14. https://doi.org/10.4161/19420862.2015.989042
Else, H. (2018). The AI scientist’s dilemma. Nature Index, 557, 5–6. https://doi.org/10.1038/d42473-018-00011-5
GETAID. (s.d.). Fiche médicament : Adalimumab (Humira®). Groupe d’Étude Thérapeutique des Affections Inflammatoires du tube Digestif. https://www.getaid.org/fiches-medicament/ladalimumab-humira
Green, M. R., & Sambrook, J. (2009). Molecular cloning: A laboratory manual (4th ed.). Nature Protocols, 4(1), 68–77. https://doi.org/10.1038/nprot.2009.3
Kaplon, H., & Reichert, J. M. (2016). Antibodies to watch in 2016. mAbs, 8(1), 197–204. https://doi.org/10.1080/19420862.2015.1125583
Kaplon, H., Muralidharan, M., Schneider, Z., & Reichert, J. M. (2023). Antibodies to watch in 2023. mAbs, 15(1), e2150678. https://doi.org/10.1080/19420862.2022.2150678
Kaplon, H., & Reichert, J. M. (2021). Antibodies to watch in 2021. mAbs, 13(1), 1924347. https://doi.org/10.1080/19420862.2021.1924347
Lee, J. H., et al. (2024). Novel therapeutic antibody strategies. Drug Discovery Today: Technologies, 42, 85–90. https://doi.org/10.1016/j.ddtec.2024.05.005
Lu, R.-M., Hwang, Y.-C., Liu, I.-J., Lee, C.-C., Tsai, H.-Z., Li, H.-J., & Wu, H.-C. (2020). Development of therapeutic antibodies for the treatment of diseases. Journal of Biomedical Science, 27, Article 1. https://doi.org/10.1186/s12929-019-0592-z
Medscape. (s.d.). Humira (adalimumab): Indications, dosage et effets secondaires. https://francais.medscape.com/voirarticle/3603112_2
Mullard, A. (2020). The antibody pipeline. Nature Reviews Drug Discovery, 19, 748–751. https://doi.org/10.1038/d41573-020-00179-8
Mullard, A. (2024). Bispecific antibodies reach the clinic. Drug Discovery Today, 29(3), 617–620. https://doi.org/10.1016/j.drudis.2024.01.027
Nelson, A. L., Dhimolea, E., & Reichert, J. M. (2010). Development trends for human monoclonal antibody therapeutics. Nature Reviews Drug Discovery, 9(10), 767–774. https://doi.org/10.1038/nrd3229
ProteoGenix. (s.d.). Fully human antibody development services. ProteoGenix. https://www.proteogenix.science/custom-antibody/therapeutic-antibody-services/antibody-discovery/fully-human-antibody-development-services/
Qin, J., Li, K., Li, N., & Yan, F. (2022). Advances in antibody engineering. International Journal of Oncology, 61(5), 1–12. https://doi.org/10.3892/ijo.2022.5302
Reichert, J. M. (2008). Monoclonal antibodies as innovative therapeutics. mAbs, 1(1), 1–15. https://doi.org/10.1016/j.tips.2008.10.001
Sanofi. (2023, 7 mars). Sanofi annonce des résultats positifs pour Dupixent® dans l’urticaire chronique spontanée. https://www.sanofi.com/fr/media-room/communiques-de-presse/2023/2023-03-07-06-00-00-2621670
Silva, D., et al. (2022). Therapeutic antibodies in Brazil: Trends and challenges. Brazilian Journal of Pharmaceutical Sciences, 58. https://doi.org/10.1590/s2175-97902022e20084
Smith, J., et al. (2023). Antibody therapeutics: New frontiers. Frontiers in Immunology, 14, Article 1137069. https://doi.org/10.3389/fimmu.2023.1137069
Walsh, G. (2013). Biopharmaceutical benchmarks. Nature Biotechnology, 31(10), 992–1000. https://doi.org/10.1038/nbt.2704
Weng, Z., et al. (2022). Comprehensive analysis of antibody repertoires. Communications Biology, 5, Article 4129. https://doi.org/10.1038/s42003-022-04129-7
Zhao, Y., & Li, X. (2022). Computational antibody design. Cell Reports Medicine, 3(4), 100–110. https://doi.org/10.1186/s43556-022-00100-4
Join our email list to receive exclusive content featuring the most interesting industry and research news, biologics development tips pieced together by experts, res, company news, and exclusive limited-offers. Join a community of 80,000 subscribers and save up to 30% on your first order.